物种形成
变种是关于物种如何形成。它是进化生物学的一个主要部分。 达尔文认为大多数物种是直接从先前存在的物种产生的。这被称为 "起源"(anagenesis):通过改变物种,或 "植物进化"。在20世纪的大部分时间里,我们认为大多数物种是由以前的物种分裂产生的:氏族发生。一般认为,大多数物种的分裂是由隔离机制造成的,或在其过程中得到帮助。 毫无疑问,曾经生活在一起的物种的物理分离是一个主要因素。许多例子说明了这一点,下面将讨论其中一些例子。 然而,过去20年的工作显示了一些其他原因。对生物的DN…
达尔文认为大多数物种是直接从先前存在的物种产生的。这被称为 "起源"(anagenesis):通过改变物种,或 "植物进化"。在20世纪的大部分时间里,我们认为大多数物种是由以前的物种分裂产生的:氏族发生。一般认为,大多数物种的分裂是由隔离机制造成的,或在其过程中得到帮助。
毫无疑问,曾经生活在一起的物种的物理分离是一个主要因素。许多例子说明了这一点,下面将讨论其中一些例子。
然而,过去20年的工作显示了一些其他原因。对生物的DNA序列的分析表明,相关物种之间往往存在一些杂交。这意味着基因已经通过这些杂交得到了转移。反过来,这意味着生殖隔离并不是物种的唯一定义,而物种的形成并不总是需要异源性(物种在生殖上的分离)。下面的章节说明了在新物种的形成中物理分离是最重要的观点。
图片库
10 图片









隔离机制
隔离机制是阻止一个物种的群体之间成功繁殖的东西。种群的生殖隔离得以建立。这对生物物种的概念特别重要,因为物种是由生殖隔离来定义的。
隔离机制可分为两组,受精前和受精后。
受精前
阻碍个体交配的因素。
- 地理上的隔离。物种出现在不同的地区,并且经常被障碍物隔开。
- 时间上的隔离。个体不交配是因为它们在不同时间活动。这可能是一天中的不同时间或不同季节。物种的交配期可能不一致。个体在其交配期或根本没有遇到对方。
- 生态隔离。个体只在其偏好的栖息地交配。它们不会遇到具有不同生态偏好的其他物种的个体。
- 行为上的隔离。不同物种的个体可能会相遇,但一个人不承认可能给出的任何性暗示。在大多数情况下,一个个体会选择自己的物种成员。
- 机械隔离。可以尝试交配,但精子的转移不会发生。这些个体可能由于大小或形态的原因而不相容。
- 配子不相容。精子转移发生了,但卵子没有受精。
受精后
妨碍交配成功的因素,如遗传不相容、杂交不育或不育。
- 雌雄同体死亡。卵子受精,但滋生体不发育。
- 杂交无生命力:杂交胚胎形成,但不具有生命力。
- 杂交不育:杂交是可行的,但产生的成体是不育的。
- 杂交种的分解。第一代(F1)杂交种是可行的和可育的,但进一步的杂交代(F2和回交)是不可行或不育的。

地理上的隔离
这被认为是最常见的物种变化的原因。第一个想到这一点的人是莫里茨-瓦格纳,一位德国探险家和自然历史学家。
瓦格纳的早期职业是一名地理学家,他出版了许多关于北非、中东和热带美洲的地理书籍。他也是一位敏锐的自然学家和收藏家,他在生物学家中最出名的就是这项工作。进化论者和生物学历史学家恩斯特-迈尔(Ernst Mayr)对瓦格纳的意义进行了阐述。p562–565
在阿尔及利亚的三年里,瓦格纳(除其他活动外)研究了不会飞的甲虫Pimelia和Melasoma。每个属都分为若干种,每个属都局限于从阿特拉斯山下到地中海的河流之间的北海岸的一段。只要过了一条河,就会出现不同的但密切相关的物种。
"......只有当少数个体越过其范围的极限边界时,才会[出现]一个新生的物种......如果没有殖民者与其他物种成员的长期持续分离,新种族的形成将永远不会成功"。
这是早期对一种地理上的变异过程的描述。1942年,迈尔重新提出了这一观点,地理上的物种演化的重要性成为进化论综合的核心思想之一。
地理上的变异的另一个术语是异地变异(allopatric speciation)。Allopatry的意思是 "不同的土地"。
空置的岛屿
火山岛的形成是没有生命的,所有的生命都必须由风或水携带到达。我们从夏威夷群岛和加拉帕戈斯群岛知道,所有的生命形式在从大陆到达岛屿时都会发生变化。
在大约6,500平方英里(17,000公里2)的范围内,夏威夷群岛拥有世界上最多样化的嗜血蝇,从雨林到山区草地都有它们的身影。已知的夏威夷果蝇种类约有800种。
研究表明,物种有明显的从老岛到新岛的 "流动"。也有殖民化回到老岛的情况,以及跳过岛屿的情况,但这些情况要少得多。
根据钾/氩放射性测年法,目前的岛屿可以追溯到40万年前(Mauna Kea)到10万年前(Necker)。夏威夷群岛中仍在海面上的最古老的成员是库雷环礁,其年代可追溯到30mya。
群岛本身是由太平洋板块在一个热点上移动产生的,它存在的时间要长得多,至少到了白垩纪。夏威夷群岛加上以前的岛屿,现在在海下,构成了夏威夷-皇帝海山链;许多水下山脉是家伙。
夏威夷的所有本地嗜血杆菌物种显然都是一个单一的祖先物种的后代,该物种在大约2000万年前殖民了这些岛屿。随后的适应性辐射是由缺乏竞争和各种各样的空位所刺激的。虽然单个怀孕的雌性有可能在一个岛上定居,但更可能是来自同一物种的群体。
夏威夷群岛上还有其他动物和植物也经历了类似的适应性辐射,尽管没有那么壮观。
环形物种
在生物学中,环状物种是一系列相连的相邻种群,每个种群都可以与隔壁的种群杂交。链的两端是重叠的。
该系列中的两个末端种群关系过于疏远,无法进行杂交。这种非繁殖--虽然在基因上有联系的'末端'种群可能在同一地区共存,从而形成一个'环'。
环状物种提供了进化的重要证据:它们说明了随着时间的推移,种群的遗传差异会发生什么。理查德-道金斯指出,环状物种 "只是在空间维度上向我们展示了在时间维度上必须始终发生的事情"。
然而,很难找到一个简单、直接的例子。
海鸥
环形物种的一个典型例子是海鸥的环极物种环。这些海鸥的活动范围在北极周围形成了一个环形,个别海鸥通常不会飞过这个环。
大体上说,海鸥可以在一定程度上与它们的邻居杂交,但在环的两端除外。
小黑背鸥和鲱鱼鸥有足够的差异,它们通常不会杂交;因此,除了在欧洲两个品系的交汇处,这组鸥类形成一个连续体。
虽然现实要复杂得多,但它很好地展示了总体思路。
恩萨蒂纳大鲵
Ensatina蝾螈是加利福尼亚中央谷地周围山区的一个环状物种。这个复合体在山脉周围形成一个马蹄形。尽管马蹄铁周围的19个种群之间可以发生杂交,但马蹄铁西端的Ensatina eschscholtzii亚种不能与东端的Ensatina klauberi杂交。这是一个说明 "几乎所有阶段的物种进化过程"(Dobzhansky)的例子。理查德-海顿认为,Ensatina是一个多物种的案例,而不是一个物种的连续体。
翠绿的莺鸟
绿莺(Phylloscopus trochiloides)有许多亚种,其中P. t . viridianus在欧洲最为熟悉。它是一个环状物种,种群在青藏高原的东部和西部出现分歧,后来在北侧相遇。它们的关系相当混乱。




同源异种
同种异构是指从一个单一的祖先物种形成两个或更多的后裔物种,都占据了相同的地理位置。这种情况现在被认为是非常普遍的。
在同种异构中,物种在居住在同一个地方时发生了分歧。经常被引用的共生物种的例子是在昆虫中发现的,它们在同一地区依赖不同的宿主植物。
作为一种物种形成的机制,同种异体的存在受到了激烈的争论。人们争论说,共生物种的证据实际上是微观地理上的物种形成的例子。一般来说,这不是现在的首选解释。相反,它是密切相关物种之间杂交的结果,随后自然选择对这种杂交的后代起作用。一个被广泛接受的同种异体的例子是东非纳布加博湖的慈鲷,这被认为是性选择的结果。
通过多倍体化进行的物种划分
多倍体引起了许多快速的物种进化事件,因为例如四倍体x二倍体交配的后代往往会产生三倍体不育的后代。
然而,并不是所有的多倍体都与它们的亲本植物进行了生殖隔离,基因流动仍然可能发生,例如通过三倍体杂交x二倍体交配产生四倍体。
许多现有的植物和大多数动物物种在其进化史上显然经历了多倍体化。成功的多倍体物种的繁殖有时是无性的,通过孤雌生殖。由于未知的原因,许多无性系生物都是多倍体。
山楂树飞
进化的一个例子是山楂蝇,Rhagoletis pomonella,它似乎正在经历同种异体的情况。
不同种群的山楂蝇以不同的水果为食。19世纪,在苹果这个非本地物种被引入后的一段时间,北美出现了一个独特的种群。这个以苹果为食的族群通常只吃苹果,不吃历史上喜欢的山楂果实。目前以山楂为食的群体通常不以苹果为食。
一些证据表明,正在发生同种异体的现象。山楂蝇比苹果蝇更晚成熟,需要更长的时间;而且几乎没有杂交的证据(研究人员已经记录了4-6%的杂交率)。
新山楂蝇的出现是进化过程中的一个例子。
杂交
罕见的是,当不同物种的个体成员交配时形成一个新的物种。通常情况下,这种杂交的产物是不孕的(不孕),或相对不孕,因此被自然选择所淘汰。
已经发现了一个成功的杂交新物种的例子。这个新物种是意大利麻雀和西班牙麻雀的杂交种,通过对其血液中的DNA进行序列分析,基本事实已经得到验证。这种鸟生活在意大利的一个地区,那里有两个亲本物种。它不与西班牙麻雀繁殖,即使它与它们并肩生活。
人工投机取巧
新的物种是由驯化的动物饲养业创造的,但最初的日期和方法并不清楚。例如,家羊是通过杂交创造出来的,不再与它们的后代Ovis orientalis产生可行的后代。
另一方面,家牛可以被认为与野牛、高尔、牦牛等几个品种是同一物种,因为它们很容易与野牛产生可育后代。
实验室物种
在实验室中创造新物种的最佳记录是在20世纪80年代末进行的。威廉-赖斯和G.W.萨特利用一个有三种不同栖息地选择的迷宫,如明/暗和湿/干,培育了果蝇,即黑腹果蝇。
每一代都被放入迷宫,从八个出口中的两个出口出来的苍蝇群体被分开,在各自的群体中相互繁殖。经过三十五代,这两组苍蝇和它们的后代在生殖上被隔离,因为它们有强烈的生境偏好:它们只在自己喜欢的区域内交配,所以不与喜欢其他区域的苍蝇交配。Rice和Hostert(1993)描述了这种尝试的历史。
黛安-多德还能够证明,在使用不同的食物类型,即淀粉和麦芽糖后,果蝇的交配偏好如何能够发展出生殖隔离。
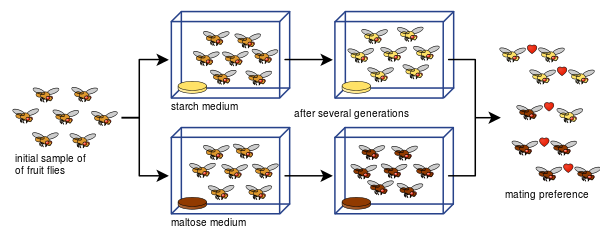
多德的实验很容易被许多其他人复制,包括用其他种类的果蝇和食物。
强化(华莱士效应)
强化是自然选择增加生殖隔离的过程。
它可能发生在同一物种的两个种群被分开后又重新接触的情况下。如果它们的生殖隔离是完全的,那么它们将已经发展成两个独立的不相容的物种。
如果它们的生殖隔离是不完全的,那么种群之间的进一步交配将产生杂种,这些杂种可能是可育的,也可能不是。如果杂交种不育,或可育但不如其祖先合适,那么就不会有进一步的生殖隔离,基本上就发生了物种形成(如马和驴)。
这背后的原因是,如果杂交后代的父母都有为自己不同环境自然选择的性状,那么杂交后代就会带有两者的性状,并且不会像父母中的任何一方那样适合任何一个生态位。杂交种的低适配性将导致选择有利于同种交配,这将减少杂交。
这有时被称为华莱士效应,因为进化论生物学家阿尔弗雷德-拉塞尔-华莱士(Alfred Russel Wallace)在19世纪末提出,这可能是物种进化的一个重要因素。
如果杂交后代比他们的祖先更适合,那么在他们接触的区域内,种群将重新合并为同一物种。
当一个物种的两种形式之间存在一个 "杂交区 "时,就需要加强分离。杂交区是分化的种群相遇和杂交的区域。杂交后代在这些区域非常常见,通常是由分化的物种进行二次接触而产生的。
如果没有强化,这两个种群或物种就会有无法控制的杂交。如上所述,在人工选择实验中可以诱导强化。
问题与解答
问:什么是变种?
答:物种演化是指物种如何形成的过程。它是进化生物学的一个主要部分。
问:达尔文对物种的形成有什么看法?
答:达尔文认为大多数物种都是直接来自于先前存在的物种,他称之为无源之水或 "植物性进化"。
问:在20世纪,人们一般如何看待物种形成?
答:在20世纪,科学家们认为大多数物种是在早期物种分裂时形成的,这被称为 "氏族发生"。人们还认为,这种分裂是由物理分离等隔离机制引起或帮助的。
问:最近的研究如何改变了我们对物种形成的理解?
答:最近的研究表明,相关物种之间的杂交可以发生,基因可以通过这些杂交进行转移。这意味着生殖隔离并不总是定义一个物种所必需的,而异源杂交(物理分离)可能并不是物种形成的必要条件。
问:物理分离与新物种的形成有什么关系?
答:曾经共存的物种之间的物理分离被认为是新物种形成的一个主要因素,科学文献中讨论的许多例子说明了这一点。
相关文章
作者
AlegsaOnline.com 物种形成 Leandro Alegsa
URL: https://zh.alegsaonline.com/art/92570
来源
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833