人類演化
人类进化是关于人类的起源。所有的人都属于同一个物种,它从非洲的发源地传播到了几乎世界所有地区。在非洲发现的化石证明了人类起源于非洲。 在这里,"人类"一词指的是智人属。然而,对人类进化的研究通常包括其他类人猿,如澳人,智人属在230万至240万年前在非洲分化(分裂)。第一代智人,也就是今天人类的祖先,是在20万年前进化而来的。 几个世纪以来,人们都知道,人类和猿类是亲戚。在内心深处,它们的解剖结构是相似的,尽管有许多表面上的差异。这就是为什么布丰和林奈,在18世纪把它们归为一个家族的原…
人类进化是关于人类的起源。所有的人都属于同一个物种,它从非洲的发源地传播到了几乎世界所有地区。在非洲发现的化石证明了人类起源于非洲。
在这里,"人类"一词指的是智人属。然而,对人类进化的研究通常包括其他类人猿,如澳人,智人属在230万至240万年前在非洲分化(分裂)。第一代智人,也就是今天人类的祖先,是在20万年前进化而来的。
几个世纪以来,人们都知道,人类和猿类是亲戚。在内心深处,它们的解剖结构是相似的,尽管有许多表面上的差异。这就是为什么布丰和林奈,在18世纪把它们归为一个家族的原因。查尔斯-达尔文的进化论说,这种基本的结构相似性来自于群体的共同起源。猿和人是近亲,属于灵长类:哺乳动物门,包括猴、猿、狐猴和塔西人。
巨猿生活在热带雨林中。人们认为,人类的进化是从一群猿类(现在被称为奥斯特罗匹克类)开始更多地生活在热带草原上开始的。稀树草原比较开阔,有树木、灌木和草。这群猿人开始用两条腿行走。他们开始用手来搬运东西。在开阔地的生活是不同的,有一个很大的优势,就是有更好的大脑。他们的大脑变大了,开始制造简单的工具。这个过程至少在500万年前就开始了。我们有两三组不同的步行猿的化石,其中一组是人类的祖先。
"人"或"男人"的生物学名称是Homo。现代人种被称为智人。"Sapiens"是"思想"的意思。智人的意思是"有思想的人"。
古人类学研究的是古人类化石、工具和其他早期人类生活的迹象。它始于19世纪,1856年发现了一个"尼安德特人"的头骨。
图片库
10 图片











智人属的直系祖先
直到20世纪20年代,非洲才发现了类人猿化石。1924年,雷蒙德-达特描述了非洲澳古猿。这个标本被称为Taung Child,是在南非Taung的一个正在开采混凝土的洞穴矿床中发现的一个澳头猿婴儿。遗体是一个保存非常完好的小头骨和一个个体头骨内部的铸件。虽然大脑很小(410厘米³),但其形状是圆形的,与黑猩猩和大猩猩的不同,更像现代人的大脑。另外,标本的犬齿很短,大孔的位置也是双足运动的证据。所有这些特征使达特确信,Taung婴儿是人类的双足祖先,是猿类和人类之间的过渡形态。
又过了20年,达特的说法才得到重视。这是在其他类似的骨架被发现之后。当时最普遍的观点是,在双足性之前,进化出了一个大脑,即两脚直立行走的能力。人们认为,类似于现代人类的智力是双足性的必要条件。事实证明这是错误的:双足性是先有的。
现在,人们认为,北极猿是现代人所属的智人属的直系祖先。穴居人和智人都是智人族的一部分,但最近的数据使人们对非洲人作为现代人直接祖先的地位产生了怀疑;它很可能是一个表亲。傲慢类原本被划分为 gracile 或 robust。健壮的Aralopithecus品种后来被重新归类为Paranthropus,尽管有些作者仍将其视为Aralopithecus的一个亚属。
在20世纪30年代,当健壮的标本首次被描述时,使用的是Paranthropus属。在20世纪60年代,健壮的品种被移入Australopithecus。最近的趋势是回到原来的分类,作为一个独立的属。


智人属
是Carolus Linnaeus选择了Homo这个名字。今天,智人属中只有一个物种:智人。还有其他物种,但它们已经灭绝了。
图中显示了其中一些人的生活地点和时间。其他一些物种可能是H.sapiens的祖先。许多很可能是我们的"表兄弟",它们是从我们的祖先线发展而来的。
人类学家仍在调查确切的世系。关于哪些应该算作单独的物种,哪些应该算作亚种,至今尚未达成共识。在某些情况下,这是因为化石很少,而在其他情况下,则是由于在对智人属的物种进行分类时使用的轻微差异。
智人属的进化主要发生在更新世。整个属的特点是使用石器,最初是粗糙的,后来变得越来越复杂。因此,在考古学和人类学中,更新世通常被称为旧石器时代或石器时代。
灵长类人
Homo habilils很可能是智人的第一个物种。它是在大约250万年前,从澳洲人发展而来的。它一直生活到大约140万年前。它的臼齿(后牙)较小,大脑比澳拉比特人大。
走向直立人
有两个拟议的物种,生活在190万年至160万年前。它们之间的关系尚未明确。其中一个叫做Homo rudolfensis。它是从肯尼亚的一个不完整的头骨中得知的。科学家认为,这只是另一个哈比利斯人,但这还没有得到证实。另一种目前被称为Homo georgicus。它来自格鲁吉亚,可能是H. habilis和H. erectus之间的中间形态,或H. erectus的一个亚种。
畸形人和直立人
直立人于1891年在印度尼西亚爪哇岛首次被发现。发现者尤金-杜波依斯最初将其称为直立人(Pithecanthropus erectus),是基于它的形态,他认为它的形态介于人类和猿类之间。直立人生活在大约180万年到7万年前。早期的标本(180万至120万年前)有时被看作是一个不同的物种,或一个亚种。 称为Homo ergaster,或Homo erectus ergaster'。
在更新世早期的1.5-1 mya,在非洲、亚洲和欧洲,据推测,Homo habilis的一些种群进化出了更大的大脑,并制造出了更精致的石器;这些差异和其他差异足以让人类学家将其归为一个新的物种--H. erectus。此外直立人是第一个真正直立行走的人类祖先。这是由锁定膝盖的进化和大孔(头骨中脊柱进入的洞)的不同位置所造成的。他们可能用火来煮肉。
一个著名的直立人例子是北京人;其他的直立人在亚洲(特别是在印度尼西亚)、非洲和欧洲都有发现。许多古人类学家现在使用Homo ergaster一词来表示这个群体的非亚洲形式。他们只将H. erectus保留给那些在亚洲地区发现的化石,这些化石符合某些要求(如骨骼和头骨),与ergaster略有不同。
尼安德塔人
尼安德特人(Homo neaderthalensis)(通常称为尼安德特人)生活在大约25万年到大约3万年前。另外,不太常见的是,作为Homo sapiens neanderthalensis:仍然有一些讨论,它是一个独立的物种Homo neanderthalensis,还是H.sapiens的一个亚种。虽然争论仍然没有定论,但线粒体DNA和Y染色体DNA测序的证据表明,新近人和H.sapiens之间很少或没有发生基因流动,因此,两者是独立的物种。1997年,时任宾夕法尼亚州立大学人类学副教授的马克-斯通金博士指出:"尼安德塔斯人和智人之间几乎没有基因流动,因此,两者是独立的物种。
"这些结果[基于从尼安德特人骨骼中提取的线粒体DNA]表明,尼安德特人没有向现代人贡献线粒体DNA......尼安德特人不是我们的祖先"。
对第二个尼安德特人DNA来源的更多调查支持了这些发现。
第三种
对在西伯利亚发现的一块指骨进行的基因分析得出了一个令人惊讶的结果。它可以追溯到大约4万年前,当时尼安德特人和现代人都生活在该地区。德国研究人员发现,它的线粒体DNA既不符合我们这个物种的DNA,也不符合尼安德特人的DNA。如果这一结果是正确的,那么这块骨头就属于以前未知的物种。DNA的差异程度表明,这个物种大约在100万年前就从我们的家族树上分裂出来了,远远早于我们这个物种和尼安德特人的分裂。
花鸟人
生活在大约10万至12万年前的Homo floresiensis因其体型小而被昵称为"霍比特人"。它的体型可能是岛屿侏儒症的结果,即大型哺乳动物在岛屿上进化出较小形态的趋势。H. floresiensis的大小和年龄都很吸引人。它是一个具体的例子,显示出与现代人类不同的衍生特征。换句话说,H.floresiensis与现代人类有着共同的祖先,但却从现代人类的血统中分裂出来,走了一条不同的进化道路。主要的发现是一副骨架,据信是一位30岁左右的女性。2003年发现的,它的年代约为18000年。活着的女人估计有一米高,脑容量只有380立方厘米,这对黑猩猩来说是很小的,还不到H.sapiens平均1400立方厘米的三分之一。
关于H. floresiensis是否确实是一个独立的物种,目前正在进行辩论。一些科学家认为,H. floresiensis是患有病理性侏儒症的现代H. sapiens。生活在发现骨架的弗洛雷斯岛上的现代人类是侏儒。这一事实与任何一种理论都是一致的。对弗洛雷斯人的一种攻击是它被发现时使用的工具只与类人猿有关。
人类抵达弗洛雷斯岛
现在在弗洛雷斯发现了石制品,其年代可以追溯到一百万年前。这些人工制品是代用品;这意味着没有人类的骨骼,但只有一种智人可以制造这些人工制品。这些文物是薄片和其他器具,共48件,其中一些有被加工出刃口的痕迹。这意味着,在那个年代,弗洛雷斯岛上就有人类存在,但它并没有告诉我们那是哪个物种。
智人
智人从大约25万年前一直生活到现在。从40万年前到中更新世的第二个温暖期,即大约25万年前,其头骨逐渐长大,并发展了以石器为基础的更复杂的技术。一种可能是发生了直立人到智人的转变。爪哇人的证据表明,直立人最初是从非洲迁出的。然后,很久以后,H. sapiens从H. erectus在非洲进一步发展。随后,非洲境内和境外的迁徙最终取代了早期的直立人。
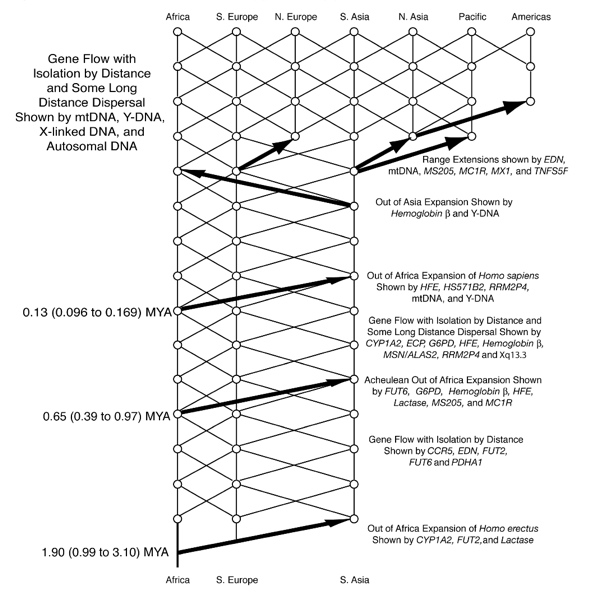
走出非洲
对人类基因组的研究,特别是对Y染色体DNA和线粒体DNA的研究,支持了近代非洲人的起源。常染色体DNA的证据也支持近代非洲起源。这个伟大传奇的细节还没有完全确定,但在大约9万年前,他们已经进入欧亚大陆和中东地区。这里是尼安德特人(Homo neanderthalensis)长期生活的地区(至少在西欧生活了50万年)。
大约在4.2万至4.4万年前,智人已经到达西欧,包括英国。在欧洲和西亚,智人在大约3.5万年前取代了尼安德特人。这一切是如何发生的细节尚不清楚。
大约在同一时间,智人到达了澳大利亚(然而最近在北领地卡卡杜的一次考古挖掘可能表明,智人在6.5万年前到达澳大利亚)。他们到达美洲的时间要晚得多,大约在1.5万年前。所有这些早期的现代人群体都是狩猎采集者。
目前的研究已经确定,人类的基因相当单一(相似)。对大多数物种来说,个人的DNA比平常更相似。这可能是由于他们相对较近的进化或托巴灾难造成的。独特的基因是由于一小部分人移居到新的环境中而产生的。这些适应性特征是智人基因组中很小的一部分,包括肤色和鼻形等外在的"种族"特征,以及在高海拔地区更有效呼吸的能力等内在特征。
H. sapiens idaltu,来自埃塞俄比亚,大约16万年前,是一个拟议的亚种。它是已知最古老的解剖学上的现代人。

物种清单
本清单按时间顺序,按属类排列。
类人猿

0.2 Mya
智人

0.6 Mya
鳆

1.9 Mya
直立派

2.8 Mya
骐骥

4.0 Mya
驼鸟
(查看-讨论)
相关网页
- ENCODE:人类基因组的完整分析。
- 古人类学
问题与解答
问:什么是人类进化?
答:人类进化是指人类的起源。它是人类从较早的物种,如澳洲人,进化到现代智人的过程。
问:人类最早出现在哪里?
答:根据在非洲发现的化石,人类首先出现在非洲。
问:智人是在多长时间前进化的?
答:第一批智人大约在20万年前进化。
问:什么是灵长类动物?
答:灵长类动物是一组哺乳动物,包括猴子、猿猴、狐猴和塔西尔。它们与人类密切相关,有许多相似的解剖特征。
问:人类的进化是什么时候开始的?
答:人类的进化至少开始于500万年前,当时有一群猿人(现在称为澳洲猿人)开始更多地生活在大草原上,并开始用两条腿走路,用手拿东西。
问:"智人 "是什么意思?
答:"人类 "或 "人 "的生物学名称是Homo。这个词来自拉丁语,意思是 "人 "或 "人类"。
问:什么是古人类学?答:古人类学研究古人类化石、工具和其他早期人类生活的迹象,以了解人类是如何随时间演变的。
相关文章
作者
AlegsaOnline.com 人類演化 Leandro Alegsa
URL: https://zh.alegsaonline.com/art/45649
来源
- ncbi.nlm.nih.gov : "Genetic support for the out-of-Africa theory of human evolution"
- cell.com : "Primate evolution – in and out of Africa"
- pnas.org : "Human origins: Out of Africa"
- anthro.palomar.edu : "Evolution's past is modern human's present"
- darwin-online.org.uk : On the Origin of Species
- ncbi.nlm.nih.gov : "Human evolution: taxonomy and paleobiology"
- doi.org : 10.1046/j.1469-7580.2000.19710019.x
- worldcat.org : 1469-7580
- pubmed.ncbi.nlm.nih.gov : 10999270
- chd.ucsd.edu : "Genomic comparisons of humans and chimpanzees"
- doi.org : 10.1146/annurev.anthro.36.081406.094339
- worldcat.org : 0084-6570
- doi.org : 10.1146/annurev-anthro-092611-145815